

Bạn có muốn tìm hiểu về code anime lands simulator Mới Nhất và cách nhập code trong trò chơi này?…

Bí mật tối quan trọng mà game thủ không thể bỏ qua – code Arena Mania mới nhất! Trên bài viết…
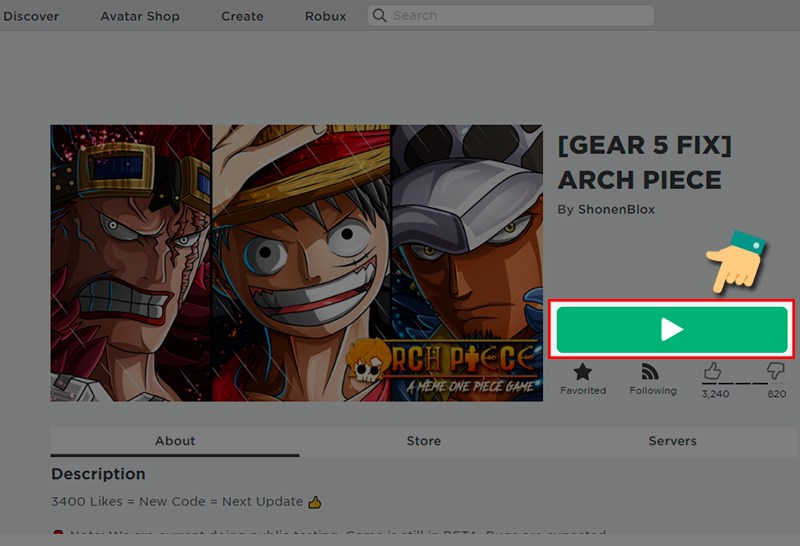
Bạn có muốn biết những thông tin mới nhất về code Arch Piece mới nhất năm 2023? Hãy đọc bài…

Bí mật về code Arcana Tactics mới nhất đang chờ đón bạn! Hãy cùng tamquoccola khám phá những mã code hấp…

Trong bài viết này, tôi sẽ chia sẻ với bạn toàn bộ danh sách mã code Arcana Roblox mới nhất…

Để tăng khả năng sức mạnh cho nhân vật trong trò chơi Arcade Hunter, tôi sẽ trình bày cho các…

Hãy cùng tôi khám phá những mã code apocalyptic rise of hero mới nhất để trải nghiệm chuỗi sự kiện…

Bạn đang tìm kiếm mã code a piece mới nhất miễn phí?.Hãy cùng tamquoccola theo dõi bài viết dưới đây…

Hãy đón chờ mã Code A One Piece game mới nhất, nơi bạn sẽ được trải nghiệm một cuộc phiêu…

Khám phá danh sách mã code Ảo Linh Sư mobile mới nhất 2023 và bí quyết nhập code để có…

Hãy khám phá danh sách các mã code Ant Legion mới nhất, với phiên bản mới nhất được tung ra…

Bạn đang tìm kiếm mã Code Anime Wrecking Simulator MỚI NHẤT? Trong bài viết này, tamquoccola sẽ cung cấp cho…

Bạn đang tìm kiếm thông tin về Code Anime World Tower Defense mới nhất? Với Anime World Tower Defense, bạn…

Bạn đang tìm kiếm các mã code Anime World mới nhất? Anime World là một trò chơi Roblox mà bạn…
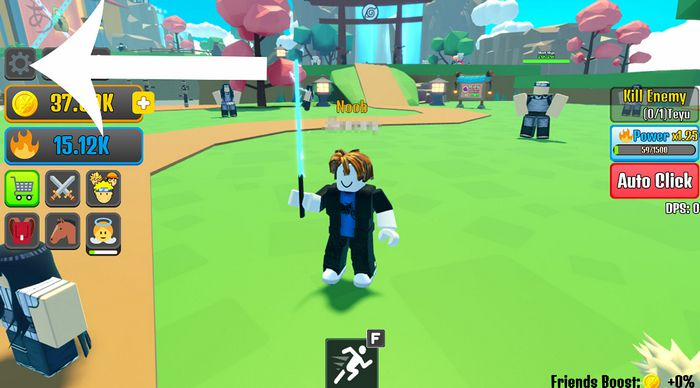
Code Anime Weapon Simulator Mới Nhất: Bạn đã từng muốn sở hữu những loại vũ khí huyền thoại trong anime…
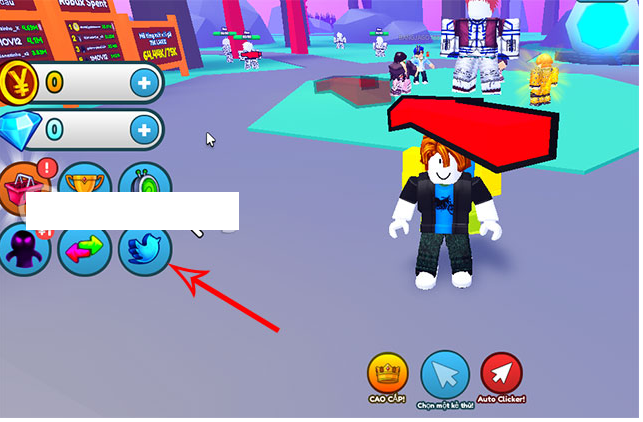
Bạn đang tìm kiếm các code Anime Warriors Simulator mới nhất? Đây là một trong những trò chơi trên Roblox…
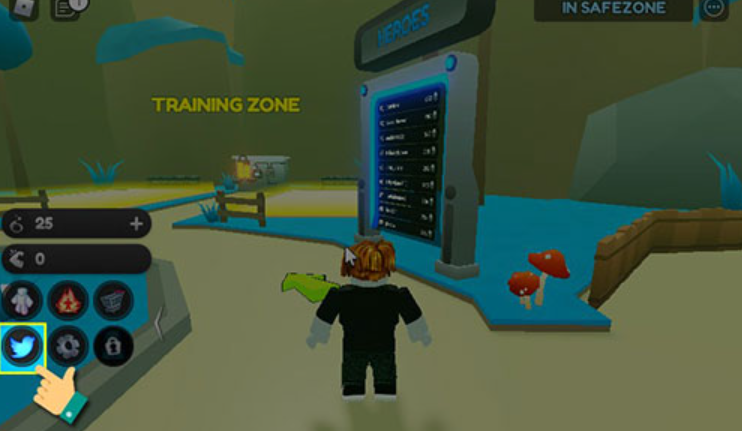
Bạn đang tìm kiếm những Code Anime Training Simulator Mới Nhất? Bạn muốn có thêm tiền vàng, gem, coin, cash,…

Code Anime Swords Simulator MỚI NHẤT – Anime Swords Simulator là một trò chơi trong Roblox, nơi bạn có thể…

Với Code Anime Story mới nhất, bạn sẽ có cơ hội nhận được các loại tiền tệ quan trọng như…

Code Anime Souls Simulator mới nhất: Anime Souls Simulator là một tựa game hành động độc đáo trên nền tảng…

Bạn muốn biết mã code Anime Showdown mới nhất? Hãy tiếp tục đọc bài viết này để cập nhật thông…

Bạn có muốn khám phá phiên bản mới nhất của code anime run beta mới nhất và biết cách nhập…

Bạn muốn khám phá mã code anime rising fighting mới nhất và cách nhập mã code cho trò chơi anime…

Bạn đã từng nghe về anime race clicker và muốn khám phá phiên bản mới nhất của trò chơi này?…

Bạn muốn khám phá code anime punching simulator mới nhất và biết cách nhập chúng? Các mã code này là…
Bạn muốn khám phá code anime power simulator mới nhất và cách nhập code anime power simulator? Anime Power Simulator…

Bạn đã từng nghe về code Anime Pet Fighters Simulator mới nhất không? Bạn muốn tìm hiểu về mã nhập…

Bạn đã từng muốn trở thành một ninja mạnh mẽ và tham gia vào cuộc chiến ninja độc đáo? Với…

Bạn đã từng nghe về code Anime Mania chưa mới nhất? Đây là trò chơi hấp dẫn dành cho các…
Bạn muốn tìm hiểu về code anime lifting simulator mới nhất và cách nhập code trong trò chơi này? Bạn…

Khi tham gia tựa game Anime Journey trong bộ game Roblox, bạn sẽ có cơ hội nhập vai thành anh…

Để nhận được Mana Multiplier bổ sung, bạn có thể đổi thẻ để mở khóa ngay lập tức những nhân…

Code Anime Hero Anh Hùng Loạn Chiến Mới Nhất là một trò chơi idle vừa được ra mắt gần đây,…

Khi bạn tham gia trò chơi Anime Guardian, bạn sẽ có cơ hội nhập vai vào nhân vật anime mà…

Mẹo nhỏ cho các game thủ yêu thích anime fruit simulator! tamquoccola đã tổng hợp dành cho các bạn danh…

Anime Fighting Simulator là một trò chơi trong hệ thống Roblox, lấy cảm hứng từ nhiều bộ anime nổi tiếng.…

Nếu bạn là một người chơi đam mê game phiêu lưu Roblox, thì bạn không thể bỏ qua code anime…

Anime Evolution Simulator là một trò chơi chiến đấu và thu thập trên nền tảng Roblox. Trong trò chơi này,…

Code anime dimensions simulator Mới Nhất là một trong những tựa game Roblox nhập vai thú vị được nhiều bạn…
Tựa game “Code anime destruction simulator Mới Nhất 2023” đang thu hút sự chú ý của cộng đồng game thủ…

Bạn có đam mê với việc tiêu diệt kẻ thù và muốn trải nghiệm những trận chiến hấp dẫn? Thì…
Hướng dẫn nhập và đổi code Anime Defense Simulator mới nhất! Tận hưởng những phần thưởng miễn phí và đá…
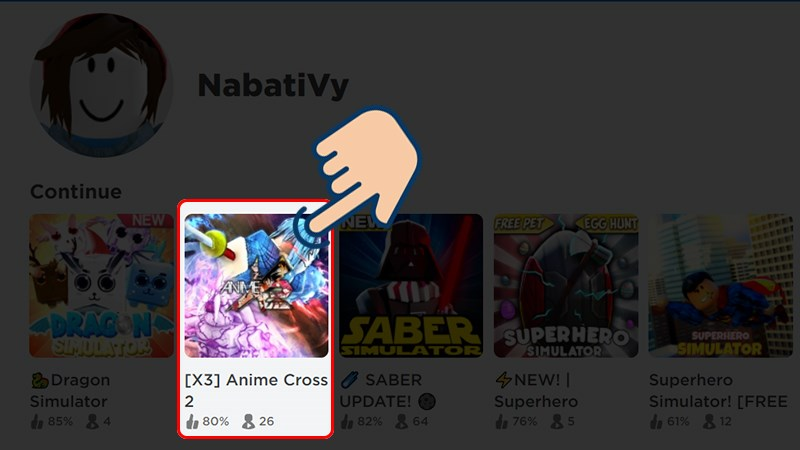
Anime Cross 2 là một trò chơi Roblox hấp dẫn, mang phong cách anime quen thuộc nhưng có lối chơi…

Hãy khám phá danh sách mã code Anime Clicker Fight mới nhất của trò chơi và cách dễ dàng nhập…
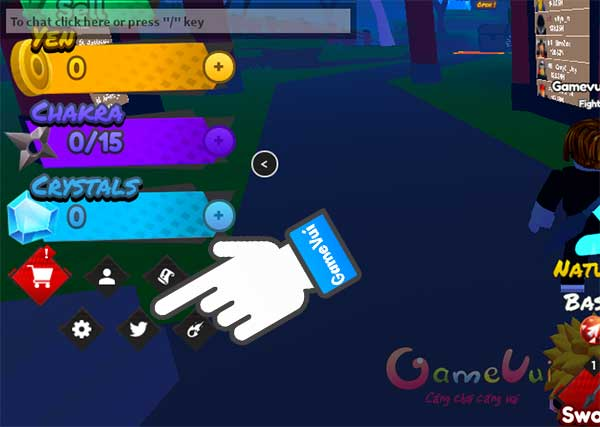
Mã code Anime Clash Simulator mới nhất và hướng dẫn sử dụng: Bạn đã chơi trò chơi click chuột trên…
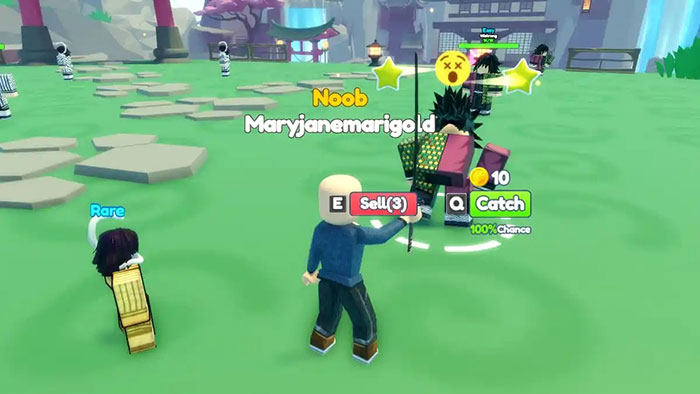
Anime Catching Simulator là một trò chơi Roblox thú vị dành cho những người yêu thích Pokémon và việc bắt…

Bạn đang tìm kiếm những cách để nâng cao khả năng chiến đấu và tích luỹ tài nguyên trong thế…
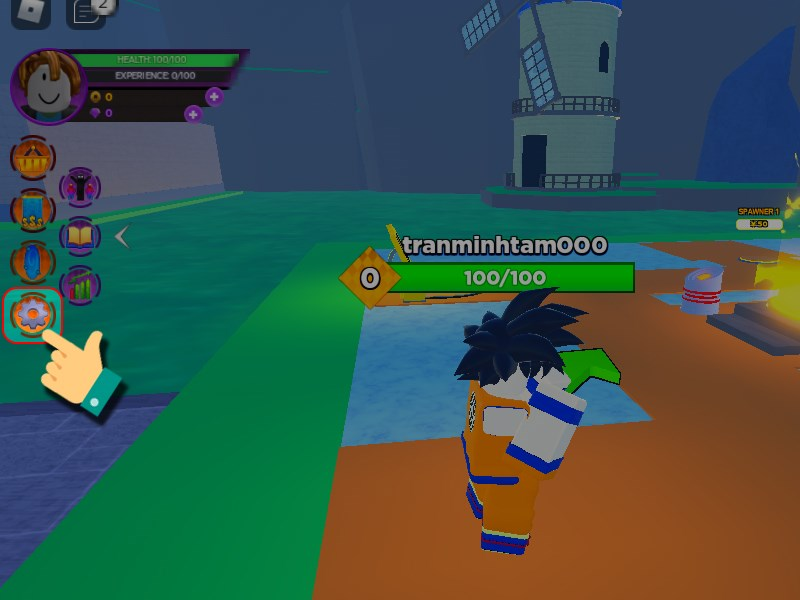
Bạn đang tìm kiếm cách tăng sức mạnh và trải nghiệm không giới hạn trong thế giới của Anime Battle…
Bạn đã nghe đến danh sách mã code Anime Battlegrounds X mới nhất chưa? Hãy khám phá cách nhập code…
Những mã code Anime Attack Simulator mới nhất giúp tăng sức mạnh, tăng tiền và tăng đá quý cho người…